
Racks & Light Shielding Panels For Rhizoboxes

Rhizobox Racks & Light Shielding Panels allow for safe, convenient placement of Rhizoboxes / Rhizonboxes and Hydroponic RootBoxes in labs, growth-rooms, greenhouses or outdoor at a standardized tilt angle - facilitating root growth along the transparent front panel and handling. Light Shielding Panels reduce the light entering the rooting zone.
Rhizobox Rack Types
Rhizobox Racks. Angled placement of Rhizoboxes / Rhizonboxes in Racks facilitates root visibility along the transparent front panel (e.g. Bonpart et al. 2020) and is required to standardize experimental conditions. VSI offers several standard / custom Rhizobox Rack designs to fit all Rhizobox dimensions and weights. The different rack types & customized designs (number of boxes, inclination angle, Shading Panels, etc.) are available via the Rhizobox configuration form (pdf). Estimate the weight of your future Rhizoboxes using the Rhizobox Calculator tool.



Standard Racks. Space between boxes allows individual boxes to be removed, optional shading panels are available to protect the rooting zone from light. 40° angled. 5 boxes per rack. Material: PP (sides) and aluminium (spacers). Comes unassembled to reduce shipping costs, all parts included. Standard Racks in the web store - in 3 sizes.



Compact Racks. NO space in-between boxes, maximize the number of root boxes that can be placed, e.g. in space-constrained growth chambers, growth rooms. The first Rhizobox per rack requires a Light Shielding Panel (see below), as the other boxes are placed front to back. 40° angled. 5 boxes per rack. Material: PP (sides) and aluminium (spacers). Comes unassembled to reduce shipping costs, all parts included. Compact Racks are available online in three sizes.



Heavy Duty Racks - Compact. Recommended for heavy boxes over ~60/70 cm in height; without space in-between boxes, 45° or custom inclination angle (fixed or adjustable), for 5-10 rhizoboxes each (depending on size). Material: aluminium (frame) / 10 mm PP (bottom grid & 1x Light Shield (in front of first box)). 4 adj. legs (to position the racks on uneven floors). Max. length 2 m. Partially pre-assembled to reduce shipping costs; all parts included & quick to install. Retrofit to other root box dimensions, if required.
Heavy Duty Racks - Single. Recommended for heavy rhizoboxes over ~60/70 cm in height. With space in-between boxes, 45° or custom angle (fixed / adjustable), vertical (90° for hydroponic RootBoxes!), for 5–8 boxes each (depending on size). Material: aluminium (frame) / 10 mm PP (bottom grid & Light Shielding Panel (in front of each Rhizobox)). 4 adjustable legs (to position the racks). Maximal length 2 m. Partially pre-assembled to min. shipping costs; all parts included & quick to install. Retrofit to other Rhizobox dimensions.
Hydroponic Rootbox Racks - Vertical. Vertical racks to place the Hydroponic Rootboxes safely and standardized.
Cooling Racks - Compact. We are currently designing and testing cooling racks for the tempered and angled placement of root boxes. Cooling Racks will be operated with standard lab coolers. Read below for Cooling Rack details.

Custom-made Rhizobox Racks. Racks & Heavy Duty and Cooling Racks can be freely configured in terms of number of Rhizoboxes, tilt angles, etc. Please contact us for a discussion and/or a custom quote based on your design.
Rhizobox Racks - Features
- Standardized tilt, increased root visibility
- Safe placement
- Different designs meet all demands (compact to removal of single boxes)
- Compatible with the use of Light Shielding Panels or Light Shields integrated
Optional
- Fitting Rhizoboxes, Rhizonboxes or hydroponic Rootboxes
- Light Shielding Panels (for standard, single racks)
Please send along: Rhizobox configuration form or contact us for custom designs.
Light Shielding / Shading Panels for Rhizoboxes

Light Shielding Panels. The shading panels reduce the light entering the rooting zone in Rhizobox Studies; read more on light effects on roots and rhizosphere. Their use is especially recommended for standard racks and heavy-duty single racks, with space between Rhizoboxes, as transparent front panels are not shaded by the back of the previous Rhizobox. Compact rack designs (see above) require at least one Light Shielding Panel (for the first Rhizobox in a row). Material: 1 mm PMMA (black). Size: According to the outer dimensions of the Rhizobox. Light Shielding Panels for standard size Rhizoboxes are available in the online shop. The panels are easily and securely attached with office binders, see example images.
Rhizobox Shading Panels - Features
- Effectively reduced Light entering the rooting zone
- Reusable
- Easy to place
- Compatible with racks



Please send along: Rhizobox configuration form or dimensions (of existing Rhizoboxes) for custom designs.
Standard Racks & Compact Racks, Light Shielding Panels for A3- A5 size Standard Rhizo(n)boxes are available in the Online Store.
Rhizobox Cooling Racks
Rhizobox Cooling Racks. We are developing novel, cost-effective cooling racks for rhizobox / rhizonbox studies. While roots normally grow in soil at a lower temperature than the air above ground, most studies place root boxes in greenhouses or climate-controlled growth rooms where the root/rhizosphere compartment has the same temperature as the shoots - for technical reasons. The higher rhizosphere temperature has consequences for growth and eco-physiological functioning of the root system, its symbionts and the soil microbiome. Therefore, we are currently testing novel, low-cost rhizobox cooling racks that will allow the temperature around the rhizoboxes to be controlled independently of the growth room (air) temperature using standard laboratory cooling equipment (i.e. circulating (cold) water in cooling coils) available in most research facilities. In addition, the boxes will have a lid system to shade the transparent front panel, allowing roots to be conveniently grown in the dark without the need to install (and remove for imaging) additional shading cloths (or commonly "aluminum foil") - allowing for time-efficient imaging. Stay tuned!
Effects of Soil Filling Procedure & Inclination on Root Visibility in Rhizoboxes
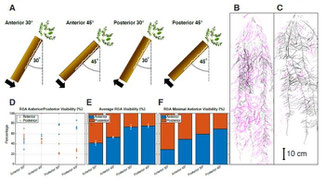
- Filling procedures and the angle of rhizobox placement affect the visibility of roots at the transparent front plate in the 3D rooting space of rhizoboxes. For example, Bonpart et al. (2020) reported that when soil was compacted against the transparent side (with the back of the boxes opened during filling) and at an inclination of 30°, only 42% of chickpea (Cicer arietinum L.) roots were observed on the front of the root boxes (see Figure). However, at an inclination of 45° (towards the front), 54% of the chickpea root system (in terms of biomass) was visible. When the soil was compacted against the back (i.e., root boxes were filled from the front), the proportion of the root system visible at the front reached approximately 73-75% at 30° and 45° inclinations, respectively. Show more
-
Bonpart and colleagues (2020) reported that the lowest anterior root system visibility was about 70% for a 45° root box inclination, while it was 59% for a 30° root box inclination. The posterior 45° condition resulted in a higher maximum anterior visibility (86%) compared to about 79% for the posterior 30° condition. In any case, filling rhizoboxes from the top (i.e., neither front nor back open) results in setups with highly variable soil densities and should be avoided. For the most comparable results, try to achieve soil densities that are common in the target ecosystem by filling similar amounts of homogenized soil into each box.
Since it is clear that the rhizobox soil filling method has an effect on root visibility, possibly due to the different soil densities at the transparent surface due to compaction, and may affect the experimental result in general, the VSI Rhizoboxes allow for convenient soil filling with either a detached front or rear panels, of from the top, to suit researcher's choice of experimental setup. Since results on the "best" angle of inclination are less clear, but larger angles to the front generally result in more visible roots, we offer rootbox racks with a standard angle of 40° (for smaller boxes) or 45° (heavy duty) - providing a compromise between maximizing root visibility and minimizing the space required to place the rhizoboxes. Racks with other angles can be made to order.
Effects of Light on Root Development
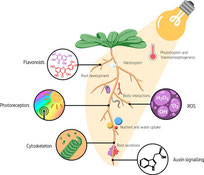
- Under natural conditions, roots grow in the dark, i.e. the absence of any illumination except for the (relatively low) light at the soil surface (organic litter layers etc.) - this simple fact has frequently being ignored in studies using transparent surfaces to study root development. For example, Zeng et al. (2019) reported that a blue light-triggered, malate-mediated photo-Fenton reaction and a canonical Fenton reaction underlie the inhibition of primary root (PR) growth induced by phosphate (Pi) deficiency in Arabidopsis.Their results also indicate that such inhibition of PR growth is not an active cellular response of Arabidopsis to Pi deficiency. Rather, it is an artifact that is induced by light when Arabidopsis seedlings are grown in transparent Petri dishes. Show more
-
Their results also indicate that such inhibition of PR growth is not an active cellular response of Arabidopsis to Pi deficiency. Rather, it is an artifact that is induced by light when Arabidopsis seedlings are grown in transparent Petri dishes. In a recent review, Canbera et al. (2022) discuss how (unintended and unwanted) experimental exposure of roots to light can bias root development and responses by altering hormonal signaling, cytoskeletal organization, reactive oxygen species (ROS), and/or flavonoid accumulation, among other things. In short, their work highlights that light exposure can alter root nutrient and/or water uptake, root responses to abiotic stresses, and root interactions with the microbiome. In addition, exposure to light can facilitate algal growth on the inner surface of transparent plates, and likely the overall composition of the microbiome during experiments.
In summary, we strongly recommend that roots be grown in the dark to get the most realistic results (i.e. relevant to soil grown plants). Therefore, we offer an (optional) Light Shielding cover for standard rhizobox racks, where the front plates are not shaded by the back of other root boxes as in compact racks (where such a cover is required at least for the first box in the sequence). In addition, we are currently working on the development and testing of cooling racks (see above) with a lid system for convenient placement of root trays in the dark. Keep the lights off for your roots!
Effects of Temperature on Root Development
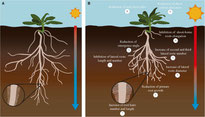
- The effect of increased temperatures on aerial parts of the plants and their responses has been well studied, whereas their influence and response on roots (and root-to-shoot signalling) has been less explored. However, it is clear that root growth & physiology are severely affected by temperature - which is commonly higher in the rooting zone of pots and rhizoboxes as opposed to ex situ studies. For example, studies in several crops have shown differential responses of aquaporins and plasmatic membrane fluidity to higher temperatures in roots (Calleja-Cabrera et al., 2020, and references therein). Similarly, nutrient balance can also be altered by changes in temperature. Show more
-
However, the effect of temperature on nutrient uptake varies greatly among crops. For example, in tomato, warmer soils limit root growth and reduce nutrient uptake, leading to a reduction in macro- and micronutrient levels (Tindall et al., 1990; Giri et al., 2017). In the grass Agrostis stolonifera, high soil temperatures result in reduced root number and increased uptake and partitioning of nitrogen, phosphorus, and potassium (Huang and Xu, 2000). In Andropogon gerardii, supra-optimal root temperatures cause a decrease in root and shoot growth. In addition, higher temperatures moderately affect nitrogen uptake, but its efficient use is severely disrupted (DeLucia et al., 1992). In contrast, warm root zone temperatures do not appear to alter N, P, and K uptake in maize to a great extent (Hussain et al., 2019). Several plant hormones involved in root development and growth have been described to mediate the temperature stress response. In particular, a role of BRs, salicylic acid (SA), ethylene (ET), abscisic acid (ABA), and cytokinin (CK) has been reported in several crops (Calleja-Cabrera et al., 2020, and references therein). There are fewer and fragmentary data on the effects of secondary metabolites on rising temperatures in roots. In maize, for example, an increase in temperature leads to a decrease in the levels of secondary metabolites such as fitosterols and terpenoids (Sun et al., 2016). Needless to say, "abnormal" root zone temperatures can also have serious consequences for root development, morphology, and overall root system architecture. Roots require an optimal temperature range for proper growth rate and function. In general, this optimal root temperature tends to be lower than the optimal shoot temperature, but roots have different optimal root temperatures depending on the species. Within this range, a higher temperature is usually associated with an altered root:shoot ratio, and a further increase in temperature would limit root development and alter root system architecture (RSA), reducing the root:shoot ratio (Ribeiro et al., 2014; Koevoets et al., 2016). RSA is defined as the organization of primary, lateral, and shoot roots. Each RSA is determined by parameters such as the length, number, and angle of these root types. RSA is the main factor that controls nutrient and water uptake efficiency because it determines the volume of soil that roots can explore in different environmental situations (Lynch, 1995). In general, exposure of roots to higher than optimal temperatures causes a decrease in primary root length, number of laterals and their angle of emergence. In addition, the increase in temperature can cause the initiation of second and third order lateral roots, which are characterized by a larger diameter (see figure for a graphical overview).
Overall, it is clear that greater efforts are needed to control the rhizosphere temperature in rhizobox experiments independently of the air temperature set in growth chambers or greenhouses. We expect that the novel, low-cost cooling racks will contribute to this effort - and help to better align the results of rhizobox studies with ex situ studies - using e.g. Rhizotron Root Windows or Minirhizotron Camera Systems for root visualisation - which are characterized by more realistic differences between soil (root) and air (shoot) temperatures. Keep your roots cool!
| References |
Rhizobox Angle, Light and Temperature Effects on Roots |
OPEN |

Rhizobox-Racks and Light Shielding Panels are accessories made to fit perfectly with our standard and custom Rhizoboxes, RhizonBoxes (for soil water sampling), and Hydroponic RootBoxes.